 脂类代谢Word文件下载.docx
脂类代谢Word文件下载.docx
- 文档编号:21923386
- 上传时间:2023-02-01
- 格式:DOCX
- 页数:38
- 大小:882.13KB
脂类代谢Word文件下载.docx
《脂类代谢Word文件下载.docx》由会员分享,可在线阅读,更多相关《脂类代谢Word文件下载.docx(38页珍藏版)》请在冰豆网上搜索。
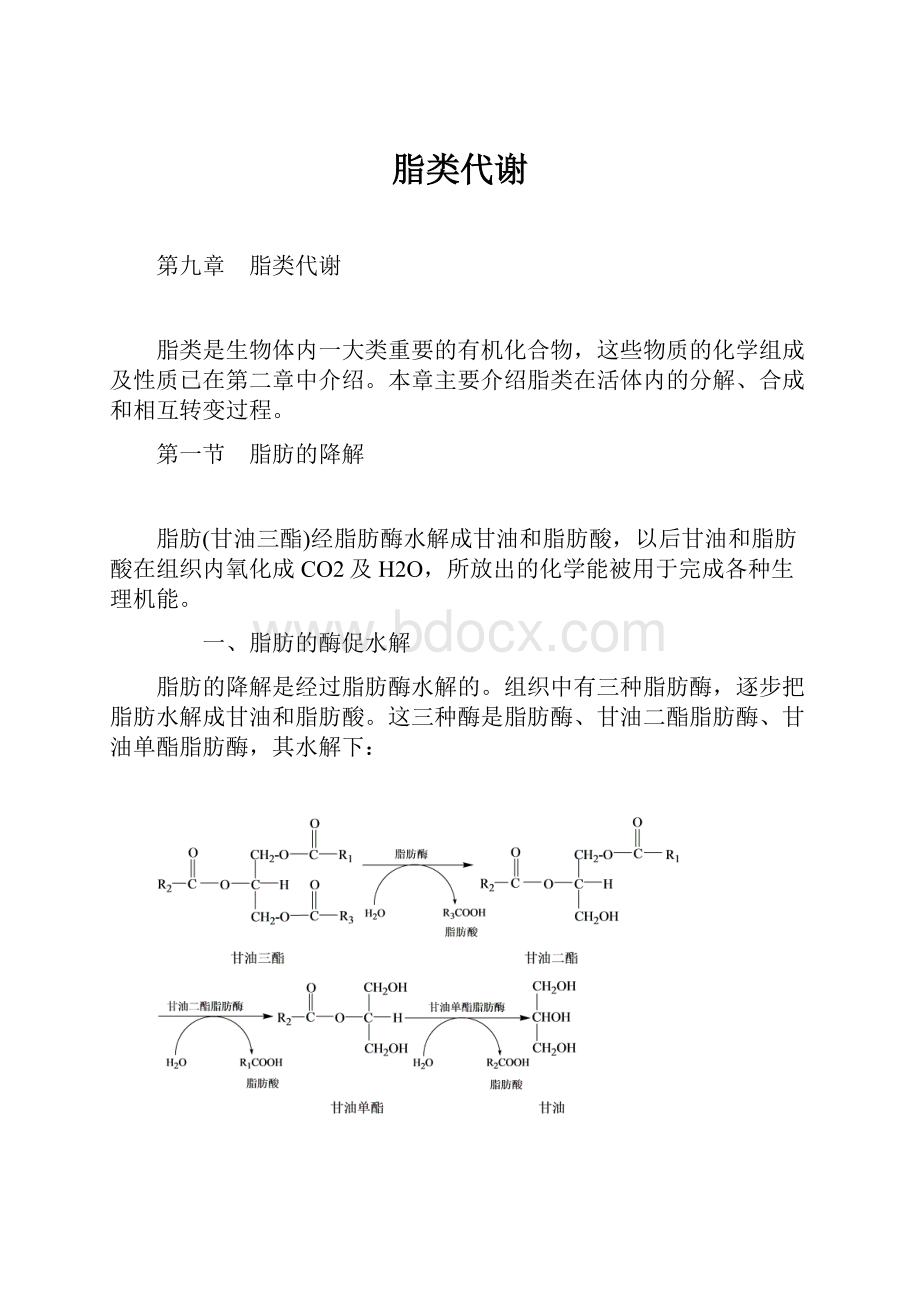
当时已知动物体缺乏降解苯环的能力,部分的苯环化合物仍保持着环的形式被排出体外。
KnooP用五种含碳原子数目不同的苯脂酸(即直链分别含l、2、3、4及5个碳原子的苯甲酸、苯乙酸、苯丙酸、苯丁酸及苯戊酸)饲养动物,收集尿液,然后分析尿中带有苯环的物质。
结果发现动物食进的苯脂酸虽然有五种,而它们的代谢产物只有苯甲酸和苯乙酸两种,苯甲酸和苯乙酸以它们的甘氨酸结合物──马尿酸和苯乙尿酸的形式从尿中排出。
换言之,动物食进的苯脂酸含有奇数碳原子(苯基的碳原子不计),则排出马尿酸,而含有偶数碳原子,则排出苯乙尿酸(表8-1)。
Knoop在上述实验的基础上提出了脂肪酸的β-氧化学说,他推论脂肪酸氧化是从羧基端的β-位碳原子开始,每次分解出一个二碳片段。
脂代谢有关酶的分离纯化、辅助因素的分析以及同位素的应用进一步阐明了脂肪酸β-氧化机制。
脂肪酸氧化的步骤如下:
(1)脂肪酸的活化脂肪酸在细胞质中首先被活化,然后再进入线粒体内氧化。
活化过程实际上就是把脂肪酸转变为脂酰辅酶A。
在细胞内有两类活化脂肪酸的酶:
(l)内质网脂酰辅酶A合成酶(acyl-CoAsynthetase)也称硫激酶(thiokinase),可活化12个碳原子以上的长链脂肪酸;
(2)线粒体脂酰辅酶A合成酶,可活化具有4~10个碳原子的中链或短链脂肪酸。
催化的反应需ATP参加,总反应式是:
该反应实际分两步进行:
首先脂肪酸的羧基与腺苷酸的磷酸基连在一起形成脂酰腺苷酸和焦磷酸,然后脂酰腺苷酸再与辅酶A化合生成脂酰辅酶A和AMP。
形成一个高能硫酯键需消耗二个高能磷酸键,反应平衡常数几乎等于1。
但由于机体内有焦磷酸酶可迅速水解反应生成的焦磷酸,成为水和无机磷,保证反应自左向右几乎不可逆地进行。
(2)脂酰辅酶A向线粒体基质转移
脂肪酸的β—氧化酶系都存在于线粒体中。
在线粒体外合成的脂酰辅酶A,中、短碳链的可以直接穿过线粒体膜进入线粒体基质中,而长碳链的不能穿过线粒体膜。
最近发现肉碱(肉毒碱carnitine)是一种载体,可将脂肪酸以脂酰基形式从线粒体膜外转运到膜内。
肉碱即L-β-羟基-γ三甲基铵基丁酸,是一个由赖氨酸衍生而成的兼性化合物。
它在线粒体膜外侧与脂酰CoA结合生成脂酰肉碱(acylcarnitine),催化该反应的酶为肉碱脂酰转移酶Ⅰ(acyl-CoAtransferaseⅠ)。
脂酰肉碱通过线粒体内膜的移位酶(translocase)穿过内膜,脂酰基与线粒体基质中的辅酶A结合,重新产生脂酰辅酶A,释放肉碱。
线粒体内膜内侧的肉碱转移酶Ⅱ(acyl-CoAtransferaseⅡ)催化此反应。
最后肉碱经移位酶协助又回到细胞质中,如图8-1。
(3)脂肪酸β-氧化作用的步骤脂酰辅酶A在线粒体基质中进行β-氧化作用。
β-氧化作用是脂肪酸在一系列酶的作用下,在α-碳原子和β-碳原子之间断裂,β-碳原子氧化成羧基,生成含2个碳原子的乙酰辅酶A和较原来少2个碳原子的脂肪酸。
β-氧化作用包括四个循环的步骤:
1
图8-1线粒体膜内外脂肪酸的转运机制
脂酰辅酶A的α-β脱氢:
脂酰辅酶A在脂酰CoA脱氢酶(acyl-CoAdehydrogenase)的催化下,在α与β碳位之间脱氢,形成反式双键的脂酰辅酶A,即α,β-反式烯脂酰CoA(Δ2反式烯脂酰辅酶A)。
在线粒体中已找到三种脂酰CoA脱氢酶,它们都是以FAD为辅基,作为氢的载体,只是分别特异催化链长为C4~C6,C6~C14,C6~C18的脂酰辅酶A。
2Δ2反式烯脂酰辅酶A的水化:
在烯脂酰辅酶A水化酶(enoyl-CoAhydratase)的催化下,反式烯脂酰辅酶A的双键上加1分子水形成L(+)β-羟脂酰辅酶A。
3L(+)β-羟脂酰辅酶A的脱氢:
经L(+)β-羟脂酰辅酶A脱氢酶[L(+)β-hydroxyac-ylCoAehydrogenase]催化,在L(+)β-羟脂酰辅酶A的C3的羟基上脱氢氧化成β-酮脂酰辅酶A。
此酶以NAD+为辅酶。
该酶虽然对底物链长短无专一性,但有明显的立体特异性,只对L-型异构体的底物有活性。
不能作用于D-型底物。
④β-酮脂酰辅酶A的硫解:
在硫解酶(thiolase)即酮脂酰硫解酶(β-ketoacyl-CoAthiolase)催化下,β-酮脂酰辅酶A被第二个辅酶A分子硫解,产生乙酰辅酶A和比原来少两个碳原子的脂酰辅酶A。
虽然β-氧化作用中四个步骤都是可逆反应,但由于硫解酶催化的硫解反应是高度放能反应,△G0/=-28.03kJ/mol。
整个反应平衡点偏向于裂解方向,难以进行逆向反应。
所以脂肪酸氧化得以继续进行。
综上所述,脂肪酸β-氧化作用有四个要点:
①脂肪酸仅需一次活化,其代价是消耗1个ATP分子的二个高能键,其活化酶在线粒体外;
②在线粒体外活化的长链脂酰CoA需经肉碱携带进入线粒体;
③所有脂肪酸β-氧化的酶都是线粒体酶;
④β-氧化过程包括脱氢、水化、再脱氢、硫解四个重复步骤。
最终1分子脂肪酸变成许多分子乙酰CoA(如图8-2)。
生成的乙酰CoA可以进入三羧酸循环,氧化成CO2及H2O,也可以参加其他合成代谢。
(4)脂肪酸β-氧化过程中的能量转变。
脂肪酸在β-氧化过程中,每形成1分子乙酰辅酶A,就使1分子FAD还原为FADH2,并使1分子NAD+还原为NADH+H+。
FADH2进入呼吸链生成2分子ATP;
NADH+H+进入呼吸链生成3分子ATP。
现以软脂酰辅酶A为例,说明其产生ATP的过程:
软脂酰辅酶A+HSCoA+FAD+NAD++H2O──→
豆蔻脂酰辅酶A+乙酰辅酶A+FADH2+NADH+H+
经过7次上述的β-氧化循环,即可将软脂酰辅酶A转变为8个分子的乙酰辅酶A。
软脂酰辅酶A+7HS-CoA+7FAD+7NAD++7H2O──→
8乙酰辅酶A+7FADH2+7NADH+7H+
每分子乙酰辅酶A进入三羧酸循环彻底氧化共形成12分子ATP,因此8分子乙酰辅酶A彻底氧化共形成8×
12=96分子ATP。
而7分子FADH2和7分子NADH进入呼吸链共产生2×
7+3×
7=35分子ATP。
所以软脂酸彻底氧化为CO2和H2O生成96+35=131分子ATP,由于软脂酸活化为软脂酰辅酶A消耗1分子ATP中的2个高能磷酸键的能量,因此净生成131-2=129个ATP高能磷酸键。
当软脂酸氧化时,自由能的变化是-9790.56kJ/mol。
ATP水解为ADP和Pi时,自由能的变化为-30.54kJ/mol。
软脂酸生物氧化净生成129个ATP,可产生30.54×
129=3939.66kJ的能量。
因此在软脂酸氧化时约有40%的能量转换成磷酸键能贮存于ATP中。
图8-2脂酰CoA的降解
2.不饱和脂肪酸的氧化
不饱和脂肪酸的氧化途径和上述饱和脂肪酸的β-氧化途径相似。
但由于它比相应的饱和脂肪酸多一个双键,所以在氧化过程中还需要有一个酶把脂肪酸分子中原有的顺式双键结构催化转变为反式结构以适于烯脂酰辅酶A水合酶的要求。
如果不饱和脂肪酸带有两个双键则还要另加一个酶把D(-)β-羟脂酰CoA催化转变成L(+)β-羟脂酰辅酶A,以适应脂酰-CoA脱氢酶的要求,使之继续按β-氧化途径进行。
前一种酶称为Δ3-顺-Δ2-反-烯脂酰CoA异构酶,它催化Δ3-顺烯脂酰辅酶A转变为Δ2-反烯脂酰辅酶A。
后一种酶称为β-羟脂酰辅酶A差向酶。
现以十八碳二烯酸为例说明上述两个酶的作用,并表示出不饱和脂肪酸的氧化途径(如图8-3)。
图8-3不饱和脂肪酸的氧化过程
3.奇数碳脂肪酸的β-氧化
生物界的脂肪酸大多数为偶数碳原子,但在许多植物、海洋生物、石油酵母等体内还有部分奇数碳脂肪酸存在。
它们按β-氧化进行,除产生乙酰辅酶A外,最后还剩下一个丙酰辅酶A。
丙酰辅酶A不能再按β-氧化继续降解,它经3步酶反应转变成琥珀酰辅酶A。
4.脂肪酸的α-氧化
Stumpf,P.K.1956年发现植物线粒体中除有β-氧化作用外,还有一种特殊的氧化途径,称为α-氧化作用。
这种氧化途径后来也在动物的脑和肝细胞中发现。
这个氧化过程是首先使α-碳原子氧化成羟基,再氧化成酮基,最后脱酸成为少一个碳的脂肪酸。
在这个氧化系统中,仅以游离脂肪酸能作为底物,而且直接涉及到分子氧,产物既可以是D-α-羟基脂肪酸,也可以是含少一个碳原子的脂肪酸。
α-氧化的机制至今尚不十分清楚,其可能的途径是:
长链脂肪酸在一定条件下可直接羟化,产生α-羟脂肪酸,再经氧化脱羧作用生成CO2和少一个碳原子的脂肪酸。
D-α-羟脂肪酸不能被脱氢酶催化,但可经脱羧和脱氢协同作用,最后产生脂肪醛。
业已证明,哺乳动物组织将绿色蔬菜的叶绿醇氧化为植烷酸后,即通过α-氧化系统将植烷酸氧化为降植烷酸和CO2。
在正常情况下,由于组织能十分迅速地降解植烷酸,所以血清中很难找到它。
但一种少见的遗传病──Refsum/S病患者,因缺少α-氧化酶系,植烷酸不被氧化。
植烷酸的α-氧化反应如图8-4。
图8-4植烷酸的α-氧化
从植烷酸的例子来看,α-氧化途径是不能彻底氧化脂肪酸的,它的作用仅仅是在β-氧化受阻时,首先进行α-氧化使β-氧化得以顺利进行。
5.脂肪酸的ω-氧化
生物体内一些中长链(如癸酸、十二碳酸等)以及少量长链脂肪酸,能首先从烃基末端碳原子即ω-碳子上被氧化生成α,ω二羧酸,称为ω-氧化。
ω-氧化涉及末端甲基的羟基化,形成一级醇,并继而氧化成醛,再转化成羧酸等步骤。
生成的二羧酸再从两端进行β-氧化。
在发现这一反应的初期,并未重视。
目前ω-氧化酶系无论从理论上或实际上已日益受到重视,其原因是可利用它来清除海水表面的大量石油。
反应过程是经浮油细菌的ω-氧化,把烃转变为脂肪酸,然后再进行脂肪酸两端的β-氧化降解。
据估计,其氧化作用速率可高达0.5g/(d·
m2),这对清除海面石油污染无疑会起重要作用。
现已从油浸土壤中分离出许多细菌,它们具有ω-氧化酶系统,可用来清除海水表面的大量浮油。
四、乙醛酸循环
许多植物、微生物中存在着一个类似于三羧酸循环的乙醛酸循环,这种循环可以看作是三羧酸循环的支路,它绕过三羧酸循环的两个脱羧反应,因此不生成CO2。
这一过程有两种关键性的酶:
(1)异柠檬酸裂解酶(isocitratelyase)将异柠檬酸分裂为琥珀酸和乙醛酸:
图8-5乙醛酸循环与三羧酸循环的关系
(2)苹果酸合成酶(malatesynthase),将乙醛酸与乙酰CoA结合成苹果酸:
苹果酸脱氢转变为草酰乙酸后,再与乙酰辅酶A结合为柠檬酸,后者再转变为异柠檬酸,于是构成一个循环反应(图8-5),这个循环反应有三个反应步骤与三羧酸循环相同,其总结果是由两个乙酰辅酶A生成1分子琥珀酸:
这样,通过一次乙醛酸循环,便有1分子的琥珀酸节余下来。
琥珀酸以后进入线粒体,可以解决因合成细胞物质而使三羧酸循环中间物减少的问题。
乙醛酸循环的另一个显著的生物学意义是在某些植物的组织中(如含油量丰富的植物种子),开辟了一条由脂肪酸转变成糖以及合成细胞物质的途径。
如果没有乙醛酸循环,脂肪酸经β-氧化产生乙酰辅酶A进入三羧酸循环后就会被完全氧化,要想合成糖则是不可能的。
但有了乙醛酸循环,就可以升高三羧酸循环中间物的浓度,为合成糖和其他物质提供前体物(如草酰乙酸沿糖异生途径合成糖,草酰乙酸和α-酮戊二酸经转氨作用生成氨基酸)。
油料作物的种子在发芽时乙醛酸循环是很活跃的。
在动物体内,因不存在乙醛酸循环,是不能将脂肪酸转变成糖的。
第二节 脂肪的生物合成
动物肝脏、脂肪组织及高等植物都能大量合成脂肪,微生物则合成较少。
合成途径是由脂酰辅酶A和L-α-磷酸甘油(3-磷酸甘油)经磷脂酸而合成的。
一、3-磷酸甘油的来源
3-磷酸甘油是合成脂肪的前体之一,它有两个来源:
一是由糖酵解中间产物——磷酸二羟丙酮在α-磷酸甘油脱氢酶(glycerolphosphatedehydrogenase)催化下,以NADH为辅酶还原形成:
二是由脂肪水解产生的甘油,在ATP参与下经甘油激酶(glycerolkinase)催化而形成。
由于脂肪组织缺乏有活性的甘油激酶,因此这种组织中三酰甘油合成所需的α-磷酸甘油来自糖代谢。
二、脂肪酸的生物合成
生物机体脂类合成是十分活跃的,特别在高等动物的肝脏、脂肪组织和乳腺中占优势。
脂肪酸的生物合成并不是其氧化降解的逆过程。
首先脂肪酸合成是在胞液中进行的,需要CO2和柠檬酸参加,而脂肪酸氧化是在线粒体中进行的;
其次脂肪酸合成酶系、酰基载体、供氢体等与脂肪酸氧化各不相同。
1.饱和脂肪酸的从头合成
(1)乙酰辅酶A的转运脂肪酸合成所需的碳源是来自乙酰辅酶A,但无论是丙酮酸脱羧、氨基酸氧化,还是从脂肪酸β-氧化产生的乙酰CoA都是在线粒体基质中,它们不能任意穿过线粒体内膜到胞液中去。
但可以通过以下途径透过膜,乙酰辅酶A与草酰乙酸结合形成柠檬酸,然后通过三羧酸载体透过膜,再由膜外柠檬酸裂解酶裂解成草酰
图8-6乙酰辅酶A从线粒体内至胞液的运转
乙酸和乙酰辅酶A。
草酰乙酸又被NADH还原成苹果酸再经氧化脱羧产生CO2、NADPH和丙酮酸,丙酮酸进入线粒体,在羧化酶催化下形成草酰乙酸,又可参加乙酰辅酶A转运循环(图8-6)。
(2)丙二酸单酰辅酶A的形成SalihWakil’s发现用细胞提取液进行脂肪酸生物合成时需要HCO3-,后来才知道乙酰辅酶A是合成脂肪酸的引物,以软脂酸为例,所需的8个乙酰辅酶A单位中,只有一个以乙酰辅酶A的形式参与合成,其余7个皆以丙二酸单酰辅酶A(malonylCoA)的形式参与合成,脂肪酸合成中,每次延长都需要丙二酸单酰辅酶A参加。
丙二酸单酰辅酶A是由乙酰辅酶A和HCO3-羧化形成的。
此反应中所用的碳原子来自比CO2活泼的HCO3—,形成的羧基是丙二酸单酰CoA的远端羧基。
催化此反应的酶为乙酰辅酶A羧化酶(acetylCoAcarboxylase),该酶的辅基为生物素(biotin)。
生物素是CO2分子的中间载体。
在大肠杆菌和植物中的乙酰辅酶A羧化酶是一个多酶复合体,可解离成三个组分:
(1)生物素羧基载体蛋白biotincarboxyl-carrierprotein,简称BCCP),相对分子质量为45000,含有两个亚基,每个亚基含一个生物素并与酶蛋白的某Lys-ε-NH2共价相连。
(2)生物素羧化酶(biotincarboxylase,简称BC),相对分子质量为98000,它也是一个含有两个亚基的酶。
(3)羧基转移酶(carboxyltransferase,简称CT),相对分子质量=130000,催化形成丙二酸单酰辅酶A。
实际上乙酰辅酶A羧化酶催化乙酰辅酶A的羧化反应分两步进行,首先是BCCP在ATP供能情况下羧化成羧化BCCP:
其次是由羧基转移酶把BCCP—CO2-的羧基转移到乙酰辅酶A的α-碳上。
乙酰辅酶A羧化酶为别构酶,又是脂肪酸合成的限速调节酶。
它有无活性的单体和有活性的聚合体两种形式,无活性的单体相对分子质量为410000,有一个HCO3-结合部位(即含有一个生物素羧基),有一个乙酰辅酶A结合部位,还有一个柠檬酸结合部位。
柠檬酸(或异柠檬酸)在无活性单体和有活性聚合体之间起调节作用。
柠檬酸有利于酶向有活性的形式转变。
缺乏它时,真核细胞乙酰辅酶A羧化酶即无活性。
柠檬酸是激活别构酶的刺激物,软脂酰辅酶A是变构抑制剂。
柠檬酸对细菌的生物合成没有调控作用。
由于丙二酰辅酶A除用于合成软脂酸外,没有其他代谢用处,所以通过调节羧化酶以调节脂肪酸的合成不会干扰其它代谢途径。
(3)丙二酸单酰辅酶A和乙酰辅酶A形成软脂酸
这是一个综合过程,要完成这循环的过程需要许多步骤,需要一个多酶复合体或多功能酶。
由于这综合反应无论在哪个有机体中都是由软脂酸合成酶催化的六步反应构成,而大肠杆菌中这个多酶复合体已经被人们纯化并进行过深入研究,故这里介绍大肠杆菌的多酶复合体。
①软脂酸合成酶多酶复合体的构成:
多酶复合体由七个成分组成,分别为酰基载体蛋白、ACP-酰基转移酶、ACP丙二酸单酰转移酶、β-酮脂酰-ACP合成酶、β-酮脂酰-ACP还原酶、烯脂酰-ACP脱水酶、烯脂酰 ACP还原酶。
它们以没有酶活性的酰基载体蛋白(acylcarrierprotein,缩写为ACP)为中心组成一簇,脂肪酸合成过程中的中间产物以共价键与载体相连。
大肠杆菌的ACP是一个含有77个氨基酸残基的热稳定性蛋白,相对分子质量为10000。
蛋白质中的丝氨酸与4-磷酸泛酰巯基乙胺(4-Phosphopantetheine)上的磷酸基团相连,这辅基也是辅酶A的组成部分(如图8-7)。
从ACP结构上看,它有一个带—SH的柔性长链,故ACP常写成ACP—SH。
由于有这个—SH,它可以从各种酰基—SCoA接受酰
基而形成
,并且释放出CoASH。
此外,由于这是长的柔性链,故它可以把酰基从一个地方传送到另一个地方,以适应多酶复合体中各种酶的需要。
各种来源的ACP的氨基酸组成十分相似。
如将不同来源的ACP交换用于不同脂肪酸合成酶系,虽然也能合成脂肪酸,但其产物因ACP而异。
图8-7磷酸泛酰巯基乙胺是CoA与酰基载体蛋白中的活性基团
②脂肪酸的生物合成程序:
(大肠杆菌中)
A原初反应。
由多酶复合物中的一个酶单体ACP-酰基转移酶(ACP-acyltransferase)催化乙酰辅酶A与ACP的-SH作用,反应如下:
乙酰基并不留在ACP上,而是转移到β-酮脂酰-ACP合成酶(β-ketoacl-ACPsynthase,以合成酶一SH或缩合酶表示)单体的半胱氨酸的-SH上,反应如下:
B丙二酸酰基的转移反应。
在ACP丙二酸单酰转移酶(ACP-malonyltransferase)催下,丙二酸单酰辅酶A与ACP-SH作用,脱掉辅酶A形成丙二酸单酰-ACP:
C缩合反应。
这一步由β-酮脂酰-ACP合成酶催化。
与酶分子中半胱氨酸-SH结合的乙酰基又转移到丙二酸单酰-ACP的丙二酸单酰基的第二个碳原子上,形成乙酰乙酰-
ACP,同时使丙二酸单酰基上的自由羧基脱羧产生CO2,反应如下:
同位素实验证明,释放的CO2的碳原子来自形成丙二酸单酰辅酶A时所羧化的HCO3-,说明羧化的碳原子并未掺入到脂肪酸中。
为什么脂肪酸的生物合成中不断地加进和失去CO2呢?
即脂肪酸合成中的缩合反应为什么是丙二酸单酰-ACP和乙酰-S-合成酶,而不是两个乙酰二碳单位呢?
原因在于羧化反应利用ATP提供能量,自由能存在于丙二酸单酰辅酶A中,当缩合反应发生时,丙二酸单酰-ACP脱羧放出大量的能量供给缩合反应所需的能量,反应过程中自由能降低,使丙二酸单酰-ACP与乙酰-S-合成酶的缩合反应比两个乙酰二碳单位分子缩合更易进行。
D第一次还原反应。
乙酰乙酰-ACP由NADPH+H+还原,形成β-羟丁酰-ACP。
催化该反应的酶为β-酮脂酰-ACP还原酶(β-ketoacylACPreductase),反应如下:
注意,这反应加氢后的产物为D型异构体,而脂肪酸氧化分解时形成的是L型异构体。
E脱水反应。
D-β-羟丁酰-ACP脱水,形成相应的α,β或△2反式丁烯酰-ACP,即巴豆酰-ACP(crotonl-s-ACP),催化该反应的酶是羟脂酰-ACP脱水酶(β-hydroxyacyl-ACPdehydrase),反应如下:
F第二次还原反应。
巴豆酰-ACP被还原为丁酰-ACP,催化该反应的酶为烯脂酰-ACP还原酶(enoyl-ACPreductase),电子供体是NADPH+H+。
在大肠杆菌和动物组织中反应如下:
丁酰-ACP的形成完成了合成软脂酰-ACP七次循环反应的第—次循环。
第二次循环是丁酰基由ACP转移到β-酮脂酰-ACP合成酶分子的-SH上,ACP又可再接受丙二酸单酰基,第二次循环即可进行。
经过七次循环后,合成的最终产物软脂酰基-ACP经硫酯酶(thioesterase)催化,形成游离的软脂酸,或由ACP转到辅酶A上,或直接形成磷脂酸。
多数生物脂肪酸从头合成只能形成软脂酸,而不能形成比它多两个碳原子的硬脂酸。
原因是β-酮脂酰-ACP合成酶对链长有专一性,它接受14碳酰基的能力很强,但不能接受16碳酰基。
可能酶与饱和脂酰基的结合位点只适合于一定的链长范围。
由乙酰-CoA合成软脂酸的总反应如下式:
8乙酰-CoA+14NADPH+14H++7ATP───→
软脂酸+8HSCoA+14NADP++7ADP+7Pi+7H2O
脂肪酸生物合成的反应程序如图8-8。
图8-8脂肪酸的生物合成过程
①乙酰CoA羧化酶②乙酰CoA-ACP转酰酶③丙二酸单酰CoA-ACP转移酶④β-酮脂酰-ACP合成酶⑤β-酮脂酰-ACP还原酶⑥β-羟脂酰-ACP脱水酶⑦烯脂酰-ACP还原酶
真核细胞的脂肪酸合成酶系与大肠杆菌不同。
许多真核细胞的多酶体系是多功能蛋白,不同的酶以共价形式连在一起,成为单一的多肽链,这有利于不同酶的协同作用。
因为含有共价键连接的多酶复合物比非共价吸附的多酶体系更稳定。
软脂酸的分解与合成途径概括起来有八点区别:
①胞内部位不同;
②酰基载体不同;
③二碳单位加入和减去的方式不同;
④电子供体或受体不同;
⑤β-羟酰基中间物的立体构型不同;
⑥对柠檬酸和HCO-的需要不同;
⑦酶体系不同;
⑧能量需求不同。
表8-2软脂酸分解与合成代谢的区别
区别点
脂酸合成
脂酸氧化
1.细胞中部位
2.酰基载体
3.二碳片段参加或断裂的形式
4.电子供体或受体
5.β-羟脂酰基中间立体异构物
6.对HCO3-和柠檬酸的需求
7.酶系
8.能量变化
细胞质
ACP
丙二酸单酰CoA
NADPH
D型
要求
7种酶,蛋白组成复合体
消耗7个ATP及14个NADPH
线粒体
CoA
乙酰CoA
FAD,NAD
L型
不要求


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- 代谢
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 1212中级汽车维修工考试试题三.docx
1212中级汽车维修工考试试题三.docx
-
333教育综合.docx
-
204届毕业生基础知识考试试题 混凝土结构设计 试题.docx
-
100以内加减运算练习题.docx
-
101软件开发工程师JAVA初级考试样卷课件word版本.docx
-
CNN代码理解.docx
-
CPA审计第4章审计抽样下载版讲解.docx
-
hr培训管理系统.docx
-
318安通科科长岗位责任制.docx
-
2044施工现场环境污染的防治措施.docx
-
12371党务平台操作手册.docx
-
Catia百格线生成宏复习过程.docx
-
725kV及以上电压等级支柱瓷绝缘子运行规范.docx
-
1144甑底链板机说明书.docx
-
100个著名初等数学问题.docx
-
201X中学寒假工作计划范文.docx
-
111 生物的特征 练习 人教版七年级上册生物.docx
-
110KV变电所设计变压器翻译.docx
-
9920第二学期学校工作总结.docx
-
0911二级技能解答.docx
-
33415设计说明书正文.docx
-
311教育学基础综合大纲.docx
-
201浙江普通高校招生选考科目考试地理试题和答案解析.docx
-
C语言程序的设计实验实验指导书及答案.docx
-
272相似三角形的性质和判定.docx
-
ACCAHA不稳定型心绞痛和非ST段抬高心肌梗死治疗指南修订版摘要.docx
-
baosteel标准对照 外标含量.docx
-
M1模拟练习题.docx
-
ARM体系课程设计实验报告.docx
-
Android面试题整理.docx
-
gaoer.docx
-
CPⅢ测设方案.docx
-
图克集装箱装车站台施工组织设计.docx
-
商业计划书.docx
-
项目类管理人员KPI绩效考核表.docx
-
完整版国产手机品牌营销策略毕业设计.docx
-
我的十年十念东方星书法创业十年工作感言吴章波.docx
-
人教版九年级英语单词听写版.docx
-
上半年陕西省初级全科主治医生考试题.docx
-
强电与弱点布线工艺及要求.docx
-
仁爱版英语七年级下册重点短语及句子.docx
-
铁路术语英文.docx
-
网络摄像机和网络视频服务器快速使用指南23.docx
-
文艺演出节目策划方案.docx
-
全国医院网址.docx
-
三大财务报表关系分析3.docx
-
上海物业服务合同版.docx
-
小学六年级英语教学工作计划范文.docx
-
社会运行理论的时空观.docx
-
苏科版九年级下第十五章《电功和电热》提优练习1及答案.docx
-
吴自选TEM8翻译真题及答案.docx
